
生命の歴史を遡るとき、私たちは幾度も「偶然」という言葉に直面する。ダーウィンの時代から現代の分子生物学に至るまで、進化とは無数のランダムな複製エラーの中から、環境に適応したものが生き残るプロセスだと定義されてきた。生命の設計図であるDNAの複製過程において、細胞内の熱的なゆらぎや化学的なノイズが偶発的なミスを引き起こす。それが突然変異の正体であり、進化の原動力であるという枠組みだ。ミシガン州立大学のリチャード・レンスキが30年以上にわたって大腸菌を7万5000世代以上培養し続けている長期進化実験は、この定説を強力に後押ししてきた。
しかし、この純粋な確率論に基づくモデルは、生命の歩みを完全に説明しきれているのだろうか。前述の長期進化実験においてすら、古典的な確率論の計算を逸脱するような劇的な適応や機能獲得が突如として現れる現象が観測されている。さらに、個体の細胞がどのようにして寿命という「時計」を共有し、老化のプロセスをこれほどまでに正確に刻んでいるのかという根源的な問いに対して、従来の生物学は十分な解答を持ち合わせていない。DNAは外部環境に対して完全に盲目であり、ひたすらにサイコロを振り続けているという前提が、今まさに限界を迎えつつある。
もし、DNAが自発的に外部の物理的な時間を計測し、進化や老化のペースを調整しているとしたらどうだろうか。
アルゼンチンの研究機関GECORPのNahuel Aquiles Garciaは、2026年3月にPLOS One誌で発表した論文[1]において、この長年の謎に対して全く新しい視座を投げかけた。DNAは宇宙空間から降り注ぐ極めて微弱な電磁信号を受信し、それを用いて自らの変異確率を調整する能動的な「量子計算機」として振る舞うという大胆な仮説である。この研究は、生命が地球という閉鎖系の中だけで自己完結しているという見方を退け、宇宙の膨張というマクロな時間軸と量子力学的なスケールで接続されている可能性を提示している。
量子力学がこじ開ける変異のマイクロタイミング
DNAの二重らせん構造は、アデニンとチミン、グアニンとシトシンという塩基対の水素結合によって繋ぎ止められている。古典的な化学の視点では、これらの結合は比較的安定したブロックの組み合わせに過ぎない。しかし、量子力学のレンズを通すと、その景色は一変する。水素結合の中核を担うプロトン(水素イオン)は、決まった位置に静止しているのではなく、「二つの谷」を持つエネルギーの地形(二重井戸ポテンシャル)の底で絶えず波のように揺らいでいる。
ここで量子トンネル効果という特異な現象が顔を出す。通常、プロトンが隣の谷へ移動するには、二つの谷を隔てるエネルギーの山を越えなければならない。しかし量子世界では、山を越える十分なエネルギーを持っていなくても、ある確率で山を「すり抜けて」反対側へ到達してしまう。このトンネル効果によってプロトンが本来とは異なる位置に移動した状態(互変異性体)が生じ、その瞬間にDNAポリメラーゼによる複製が重なると、細胞は誤った塩基を組み込んでしまう。これが突然変異の微視的な発端となる。
Garciaの研究が踏み込んだのは、このトンネル効果が発生するマイクロタイミングである。変異が定着するかどうかは、エラーを検知する修復システムの作動と、不自然な結合が解消されるまでの時間的なレースに依存する。もし外部からの何らかの物理的な介入によって、このプロトンが反対側の谷に滞在する時間がわずかでも延長されたり、トンネル効果の発生確率が偏らされたりすれば、結果として突然変異の頻度は操作されることになる。細胞の老化という現象も、この微小なエラーの蓄積の産物である。時間とともに進行するミスの蓄積ペースが変われば、生物学的な時計の進み方もまた変化する。
宇宙の膨張を聴き取るアンテナの構造
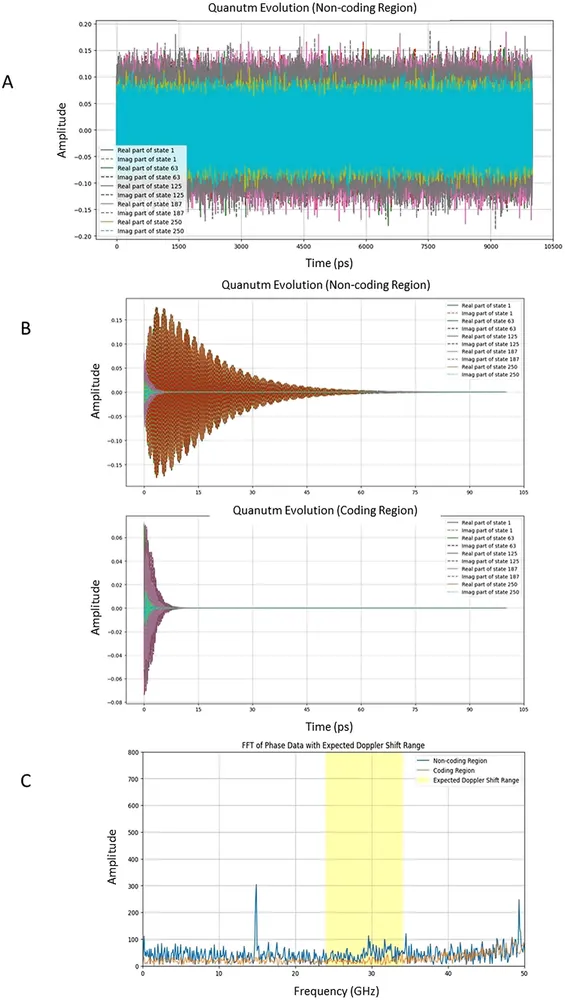
問題は、何がその介入を引き起こすのかという点にある。Garciaが着目したのは、宇宙の誕生であるビッグバンの名残として今も全宇宙を満たしている宇宙マイクロ波背景放射(CMB)などの環境電磁波である。DNAの階層的に折り畳まれた分子構造は、特定の周波数の電磁波と相互作用するフラクタルアンテナの特性を持つことが先行研究で示唆されていた。実際に過去の研究では、水溶液中のDNAが34GHz付近のマイクロ波に対して共鳴応答を示すことが報告されている。
宇宙は絶えず膨張を続けているため、遠方から届くCMBの波長は引き伸ばされ、地球上の観測者から見ればごくわずかな周波数の低下(ドップラーシフト)が生じる。論文のシミュレーションでは、34GHzの周波数帯域において、3時間で0.0008Hzという極めて微細な周波数シフトを想定した。このドップラーシフトによる波長の伸びこそが、宇宙の時間の流れを客観的に示す指標となる。
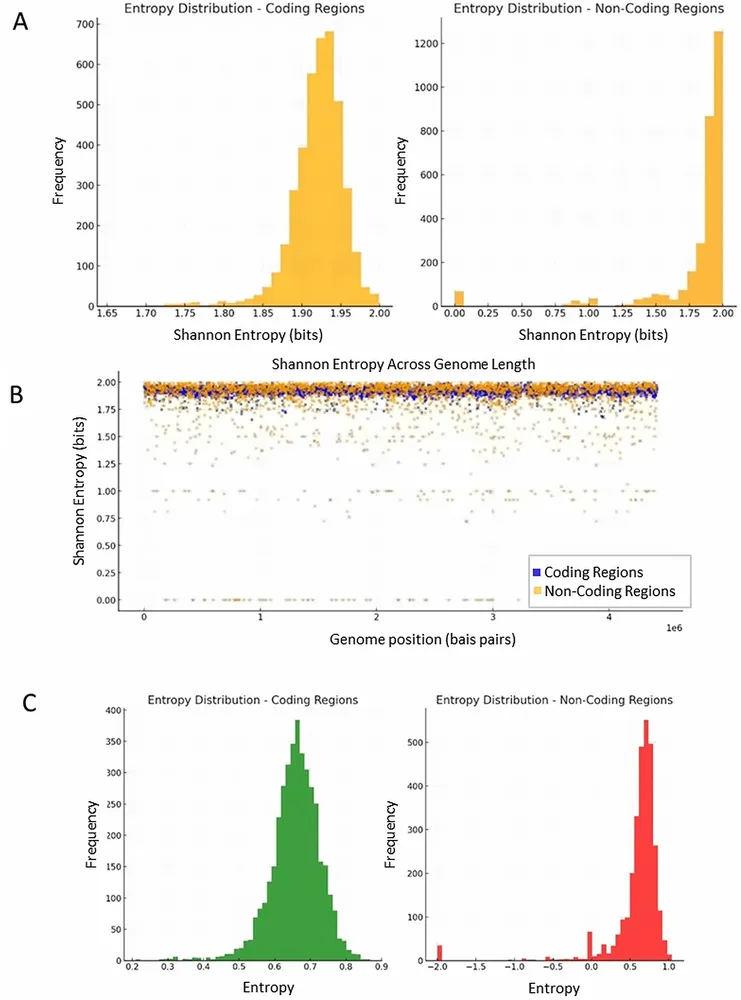
Garciaは、結核菌(Mycobacterium tuberculosis)のゲノム配列を用いて、この微弱な宇宙の信号がDNAの量子状態にどのような影響を与えるかをコンピュータ上でシミュレートした。DNAの塩基配列を二値の量子状態にマッピングし、シュレーディンガー方程式を用いて時間発展を追跡したのである。ここで彼が焦点を当てたのは、タンパク質の設計図となる「コーディング領域」と、これまで明確な役割を持たないジャンクDNAと呼ばれてきた「非コーディング領域」の明確な違いである。
解析の結果、両者の情報論的な複雑さを示すシャノンエントロピーに明確な差が確認された。結核菌ゲノム全体のうち約8.92%を占める非コーディング領域は平均1.81ビット、コーディング領域は平均1.92ビットであり、その差は統計学的に極めて有意(p=5.03×10^-69)であった。さらに、量子状態の不確実性を示すフォン・ノイマンエントロピーにおいても、コーディング領域(平均0.6576)が低分散で安定しているのに対し、非コーディング領域(平均0.5925)は分散が大きく、外部からの影響を受けやすい揺らぎを持っていた。
この数値は、DNAという単一の分子内に、二つの異なる役割を持つ構造が同居していることを物語る。非コーディング領域は、外部からの微弱な電磁波の変動を敏感に感知するアンテナ兼センサーを担い、そこで捉えられた宇宙の時間の情報は、量子もつれ(エンタングルメント)を介して安定した演算回路であるコーディング領域へと転送される。こうしてDNA全体が一つの量子システムとして連動し、細胞の生物学的なリズムを外部環境と同調させているという構図である。これは、量子鍵配送(QKD)において、高いエントロピーを持つ領域を介して安全に情報が伝達される仕組みを彷彿とさせる。
| 比較項目 | 従来の突然変異モデル | 本研究が提示する量子モデル |
|---|---|---|
| 変異の原因 | ポリメラーゼの確率的エラーや熱ゆらぎ | 外部電磁波によって変調されるプロトントンネリング |
| DNAの役割 | 情報の保存媒体となる化学的なテープ | センサーと回路を持つ精巧な量子システム |
| 進化の構造 | 完全なランダム性に依存し自然選択が働く | 外部からの情報入力によって変異確率が微調整される |
| ジャンク配列 | 進化の過程で蓄積した意味を持たない残滓 | 環境変動を捉え量子干渉を維持する適応型受容器 |
確率共鳴が繋ぐマクロの波とミクロの揺らぎ
ここで一つの大きな物理学的障壁が立ちはだかる。CMBが持つエネルギーはあまりにも微小なのだ。計算上、一つのヌクレオチドがCMBから受け取るエネルギーは1秒間に約10^-32ジュールに過ぎない。プロトンがエネルギーの山を越える(あるいはトンネル効果のトリガーとなる)ために必要な活性化エネルギーは約0.1電子ボルト(1.6×10^-20ジュール)であり、DNAがこのエネルギーを単独で蓄積するには、何百万もの塩基が協力したとしても数日を要してしまう。
このエネルギーの圧倒的な不足を補い、マクロな宇宙の波とミクロなDNAの挙動をリンクさせる鍵が「確率共鳴(Stochastic Resonance)」という現象である。
確率共鳴とは、特定の微弱な信号が単独では閾値を越えられなくても、背景にランダムな雑音が存在することで、その雑音に乗せられる形で信号が浮き上がり、システムに影響を与える現象を指す。騒がしいレストランの中で、遠くの席で発せられた自分の名前だけが、周囲のノイズに押し上げられてふと耳に届くような感覚に近い。
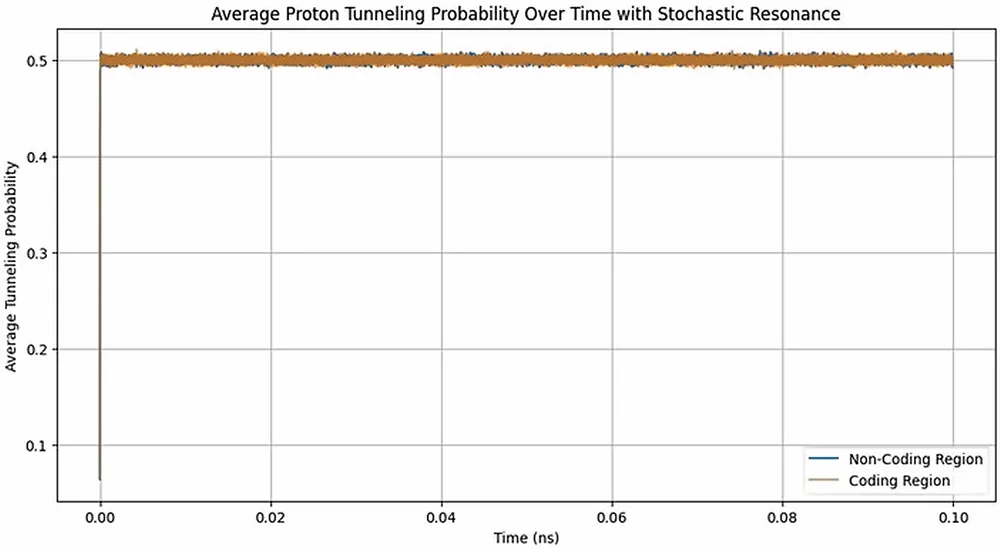
細胞内は常に37度前後の熱ノイズと生化学的な分子の衝突という荒波に満ちている。Garciaのモデルでは、DNAはこの細胞内の熱エネルギーを主動力としてプロトンの移動を引き起こしている。しかし、その荒波の上に、ドップラーシフトを伴うCMBの規則的な微波が乗ることで、プロトンがどちらの谷に落ちるかという微視的な確率が偏るのである。
実際に、外部からの34GHzの微弱な摂動とノイズを組み合わせたシミュレーション(シミュレーション時間は0.1ナノ秒)では、実際の結核菌ゲノムを用いた場合においてのみ、プロトンのトンネリング確率に明確な変化(p値約0.004)が見られた。一方、塩基の配列を無作為に並べ替え、コーディング領域と非コーディング領域の生物学的な構造を破壊したコントロール配列では、この有意な変化は消失した。DNAの塩基配列そのものが、宇宙からのノイズを読み解き、意味のある信号として抽出するための物理的なフィルターを備えていることを示している。
未知の領域への実験的挑戦と生命科学のパラダイムシフト
この研究は、生命現象の根底に宇宙論的なスケールの物理法則が関与している可能性を提示した点で極めて野心的である。加えて、ヒトの概日リズムを制御するCRY1遺伝子を用いた追加解析でも結核菌と同様の量子的な振る舞いが確認されており、このメカニズムが種を超えて保存された普遍的なシステムである可能性を示唆している。
一方で、著者のGarcia自身も論文内で強調しているように、現段階ではこれはあくまで理論的なトイモデルの域を出ない。実際の細胞内におけるDNAの塩基対は、周囲の水分やタンパク質との相互作用によって非対称で複雑なポテンシャルエネルギーを持っており、本シミュレーションで用いられた対称的な二重井戸モデルは現実の物理環境を大胆に抽象化したものである。

また、宇宙空間から届くGHz帯の電磁波は、地球の大気や生物の組織を通過する過程で大きく減衰し、さらに人工的な電波や熱放射のノイズにかき消されてしまう。CMBの放射が直接的に生体内のDNAを駆動しているというよりは、DNAの分子構造が持つ時間依存的な微小信号に対する感受性を検証するための思考実験の題材としてCMBが選ばれた、と解釈するのが科学的に誠実な態度である。
この壮大な仮説を現実の生物学に定着させるためには、生体内での実験的な裏付けが不可欠となる。Garciaは具体的な検証アプローチとして、外部の電磁場を完全に遮断するファラデーケージの中で大腸菌を長期培養し、リアルタイムで突然変異の発生頻度やパターンを追跡する実験を提案している。もし外部電磁波の遮断によって変異のダイナミクスに有意な変化が生じた場合、あるいはCRISPRを用いて非コーディング領域のアンテナ配列を編集することで突然変異のパターンが制御できた場合、この量子パラダイムは決定的な裏付けを得ることになる。
もしこの仮説が実証されれば、その波及効果は学術界にとどまらない。産業や医療における応用可能性は計り知れない広がりを見せる。たとえば、特定の周波数の微弱な電磁場を人為的にコントロールすることで、DNAのプロトントンネリング確率を操作できるとすれば、がん細胞における体細胞変異の発生を抑制したり、加齢に伴う細胞の老化プロセスを人為的に遅延させたりする全く新しい物理的アプローチの医療が誕生する。また、合成生物学の分野においては、酵素や微生物の進化プロセスをランダムな変異に頼らず、電磁場を用いて特定の適応方向へ誘導・加速させる技術への展開も想定される。
私たちがランダムと呼んできた進化のサイコロは、もしかすると宇宙の膨張という途方もなく巨大な時計の歯車と噛み合っているのかもしれない。DNAが星々の歴史を読み解き、進化の歩みを自ら刻む「宇宙の受信機」でもあるという視座は、私たちと宇宙との境界線を静かに、そして根本的に溶かし始めている。


